Santiago Ramón y Cajal1852–1934
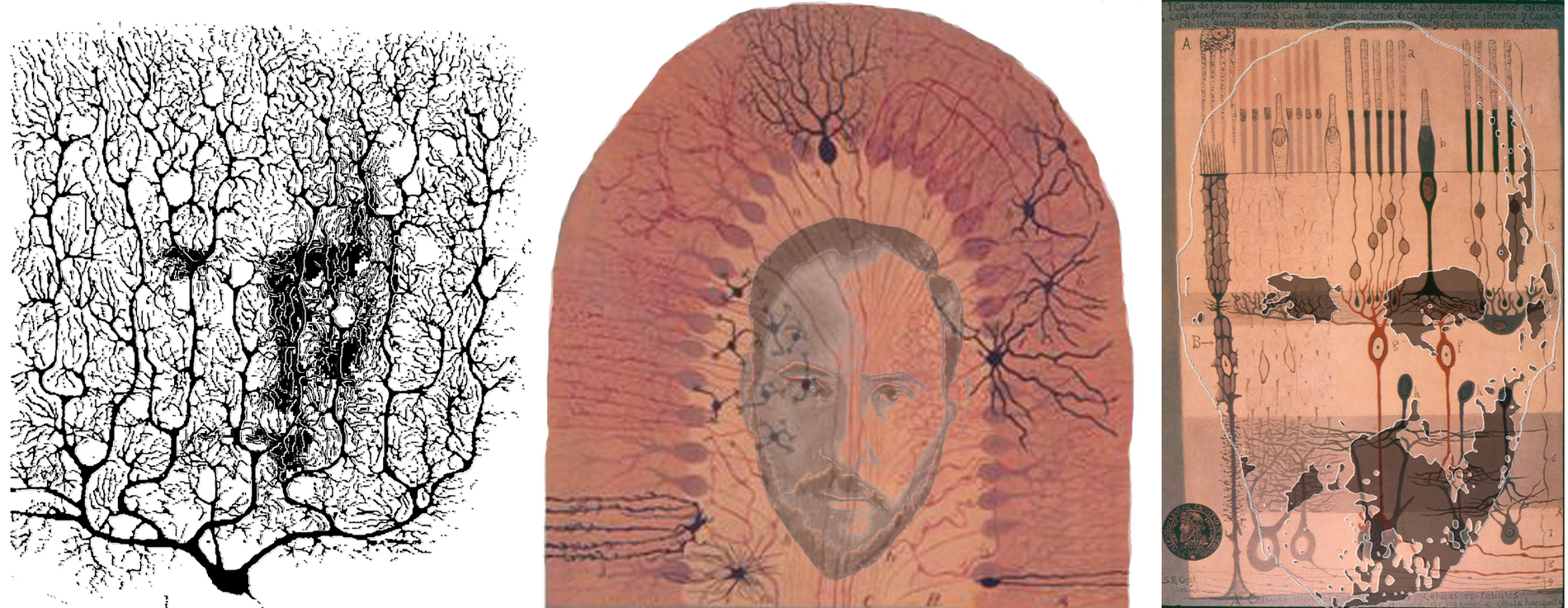
Golgi’s black reaction transformed histological studies of the nervous system. After having been first exposed to the process in 1887, Cajal adopted and adapted it, and in the next year he published the results of his initial investigations in the cerebellum and in the retina. In the cerebellum he was able to establish the individuality of the cellular elements and to recognize the presence of an ordered pattern of connectivity. Of particular relevance for his further research were the connections involving the Purkinje cells, at both their rich dendritic region and at the soma – with the arborizations of climbing fibres and with the basket-like terminations of the cell or the molecular layer. Purkinje cells, and the cerebellar structure, played an important part in the fierce debate that was raging about the structure of the nervous system. On the one hand, the ‘reticularists’ (like Golgi) argued that the nervous system was a single interconnected and continuous network; nerve cells were not independent units. On the other hand, the nerve cells (or neurons, as Waldeyer called them in 1891) were considered to be independent units which communicated by electrical signals along well defined paths and in a ‘dynamically polarized’ way: the signals passed from the dendro-somatic region of one cell via its axon terminal to the dendro-somatic region of the next cell. Cajal started elaborating this view after his first studies applying the Golgi technique to the cerebellum, and had to defend it for much of his scientific life. The retina, the second structure studied by Cajal, would be seen by him as providing strong evidence in favour of the neuron doctrine. In particular it gave him important clues for the identification of the direction of signal transmission at the interneuronal contacts. This was because of its relatively simple structure and of the evident direction of visual signal flow from photoreceptors toward the ganglion cells. However, it was Tartuferi, one of Golgi’s students, who first applied the silver staining technique to the retina in 1887. When Cajal viewed the same structure he did not represent it in the same way. He took the principal distinction between the rods and cones to support Schultze’s duplicity theory. He also sought to retain the distinction for the fibres in connection with the receptors. In this regard he did find different bipolars for rods and cones in some species. The presence of lateral connections from the horizontal and amacrine cells presented a problem for Cajal’s conception of the manner in which vision operated, and he tended to exclude these two classes of cells from the operative visual network of the retina, by assigning them ‘energetic’ or modulator roles. He considered that there was a point-by-point analysis of the retinal image, with the punctate pattern being retained at central sites. Lateral connections in the visual pathways could disturb this pattern of projection, and Cajal sought to see in his histology the evidence for such a sequence. It would seem that he saw what he sought by matching the microscopic appearances to his operational conception of retinal function. The functional paradigm of retinal and visual physiology adopted by Cajal was clearly inspired to the process of photography. Not only did he develop great skills as a photographer but he also applied them to stereoscopic vision. The technique he devised is not unlike the principle of random dot stereograms and had the intention of encrypting messages. Cajal is shown in a Purkinje cell stained by Golgi’s black reaction, crowned by the cells of the cerebellum, and in his ‘generous retina’.